Bild 1 Schematischer Querschnitt durch die Netzhaut
Künstliche Systeme zur Aufnahme und Verarbeitung von Bilddaten sind bis heute viel weniger leistungsfählig als biologische. Dies ist insbesondere in natürlichen Umgebungen augenfällig, auf die biologische Systeme infolge ihrer langen Entwicklungszeit optimiert sind. Deshalb überrascht es kaum, dass nicht nur Naturwissenschafter, sondern auch Ingenieure schon vor geraumer Zeit begonnen haben, sich mit biologischen Methoden auseinanderzusetzen. Als vorderstes Glied in der Kette der visuellen Datenverarbeitung hat das Auge natürlich die Aufmerksamkeit zuerst auf sich gelenkt. Die ältesten und am weitesten verbreiteten künstlichen Abbildungsmethoden sind derjenigen des menschlichen Auges prinzipiell sehr ähnlich und mittlerweile durch jahrhundertelange Entwicklung sogar überlegen. Bei der Bildaufnahme jedoch, das heisst bei der Umwandlung von elektromagnetischen Schwingungen in ein elektrisches Signal. können die heutigen Standardsysteme (elektronische Kameras) in vielen Belangen nicht mehr mithalten. Und schon für die ersten Bildverarbeitungsschritt wird typischerweise ein Computer zur Hilfe genommen, der Sekunden bis Minuten braucht, um Bildverarbeitungsoperationen durchzuführen, die von der Netzhaut mit minimalem Leistungsaufwand in einigen Millisekunden realisiert werden, obwohl die biologischen Schaltkreise etwa eine Million Mal langsarner sind als die eines Computers und zudem eine erhebliche statistische Variabilität aufweisen. Aus diesem Grunde ist in den letzten Jahrzehnten die Retinal als Bildaufnahmeund -vorverarbeitungssystem vermehrt ins Zentrum des Interesses gerückt..
Simulation oder Emulation?
Die Ursache für diese gewaltige Diskrepanz
zwischen rechnergestützter und biologischer Bildverarbeitung liegt
hauptsächlich darin, dass - im Gegensatz zu Abbildung und Bildaufnahme
- Struktur und Datenrepräsentation solcher Systeme grundverschieden
sind. Anders als ein computerbasiertes System ist die Netzhaut topographisch
dem Bild angepasst, das sie aufnimmt. Die Bildverarbeitungsoperationen,
die die Netzhaut ausführt, benötigen nur eine lokale Vernetzung
der Datenverarbeitungselemente und ermöglichen deswegen kurze Signalwege
und somit eine schnelle parallele Datenverarbeitung auf kleinstem Raum.
Voraussetzung dafür ist, dass die Datenverarbeitung und -zwischenspeicherung
direkt am Ort der Bildaufnahme geshieht. Ein System, das aus einer Videokamera
und einem Computer besteht, kann diese Voraussetzung nicht erfüllen,
da Aufnahme, Speicherung und Verarbeltung der Daten konzeptuell und räumlich
getrennt sind.
Bildsignale werden als analoge physikalische Grössen von einem Sensor empfangen und in ebensolche Grössen transformiert. Innerhalb der Retina wird diese Reprräsentationsform für Signale und gespeicherte Werte beibehalten, während ein konventioneller Prozessor die Sensorsignale erst in digitaler Form verarbeiten kann. Durch die analoge Datenverarbeltung der Netzhaut können physikalische Prozesse viel direkter und deshalb effizienter ausgenutzt werden als von einem digitalen Prozessor, dessen ineffiziente Kodierungen sich natürlich in Grösse Leistungsverbrauch und Komplexiät der Schaltungen niederschlagen.
Die Reproduktion der Beziehung zwischen einem Eingangs- und Ausgangssignal eines Systems durch ein anderes System wird Stimulation genannt. Ein System aus Videokamera und Computer kann also zum Beispiel dazu verwendet werden, die Netzhaut zu simulieren. Wenn aber das ursprüngliche System auf dem Niveau der einzelnen Datenverarbeltungselemente kopiert wird, also ein struktureller und funktioneller Isomorphismus besteht, wird dies Emulation genannt. Aufgrund der vorangehenden Betrachtungen können wir also annehmen, dass eine Emulation der Netzhaut effizienter ist als ein Simulation. Eine Emulation setzt allerdings voraus, dass man die Funktionsweise des ursprünglichen Systems versteht und nicht nur seine Übertragungsfunktlon kennt. Bei der Netzhaut ist dies im Gegensatz zu den meisten anderen Hirnregionen weitgehrnd der Fall.
Aufbau der Retina
Dass die Netzhaut zu den am besten verstandenen
Teilen des Hirns gehört, liegt in erster Linie daran, dass die sich
relativ gut untersuchen lässt: Sie ist peripher angeordnet und daher
leicht zugänglich für anatomische und physiologische Analysen.
Zudem
besteht sie nur aus einer kleinen Anzahl lokal vernetzter Zelltypen, die
von anderen Hirnregionen klar abgegrenzt sind. Diese Eigenschaften und
die Tatsache, dass typische Eingangssignale gut bekannt und kontrollierbar
sind, erleichtern die Untersuchung der Retina.
Die Retinae der meisten Wirbeltiere sind sicc sehr ähnlich in Struktur und Signalcharakteristik. Sir enthalten fünf Hauptzelltypen, die in drei Schichten angeordnet sind (Bild 1). Die äussere Schieht besteht aus Photorezeptorzellen, die Lichtreize in elektrische Signale verwandeln, die mittlere aus Bipolar-, Horizontal- und Amakrinzellen, die die Signale analog verarbeiten, und die innere Schicht aus Ganglienzeilen, deren Ausgangssignale als Impulse durch den Sehnerv in die höheron Zentren der Signalverarbeitung weitergeleitet werden. Die Photorezeptoren sind auf der dem Lichteinfall abgewandten Seite der Netzbaut angeordnet; die Retina wird also von der Rückseite her beleuchtet.
Es werden zwei Typen von Photorezeptorzellen unterschieden. Die Zapfenzellen sind über die ganze Netzhaut verteilt. In der Fovea centralis, einem Bereich im Zentrum der Netzhaut mit einrm Durchmesser von etwa einem halben Millimeter, ist dieser Zelltyp mit einer besonders grossen Dichte vertreten. Der Zellabstand von etwa 2.5 mm in der Fovea centralis limitiert unsere maximale Sehauflösung auf etwa eine halbe Bogenminute. Wegen der geringeren Zapfendichte am Rand der Netzhaut verschlechtert sich die Sehauflösung in der Peripherie des Sehfeldes auf etwa ein Grad.. Die Zapfenzellen sprechen auf Reize im Tageslichtbereich an. Sie können in drei Klassen eingeteilt werden, die in verschiedenen - teilweise sich überlappenden - Spektralbereichen empfindlich sind mit mittleren Wellenlängen, die blauem, grünen und orangem Licht entsprechten.
Die Stäbchenzellen, die sich beim Menschen auf der ganzen Retina ausser in der Fovea finden, haben ein breitbandigere spektrale Empfindlichkeit. Sie kommen im Dämmerlicht und nachts zum Einsatz und sättigen bei starker Beleuchtung. Stäbchen sind so lichtempfinlich, dass sie fast einzelne Photonen detekterien können. Die Netzhaut hat etwa 125 Millionen Photorezeptoren, von denen die überwiegende Mehrheit Stäbchen sind. Photorezeptoren wandeln eine Änderung in der einfallenden Lichtintenzität logarithmisch in eine Änderung ihres Ansgangspotentials um. Sie haben eine relativ kurze Ansprechzeit und zeigen eine langsame Adaptierung. Ein Photorezeptor ist über etwa fünf Grössenordnungen in der Beleuchtungsstärke empfindlich; mit Zapfen und Stäbchen zusammen deckt das Auge also etwa zehn Dekaden ab, von Sternenlicht bis zu gleissendem Sonnenlicht. Im Gegensatz zu fast allen anderen Nervenzellen, die auf einen Reiz mit Depolarisation und Ausschüttung von Neurotransmitter an chemischen Synapsen reagieren, werden Photorezeptoren in Wirbeltieren durch Stimulation hyperpolarisiert. Das bedeutet, dass sie in der Dunkelheit aktiv sind und diese Aktivität bei Beleuchtung abnimmt. Benachbarte Photorezeptoren können durch elektrisch resistive Verbindungen (Gap junctions) miteinander gekoppelt sein, so dass eine lokale Mittelung des Signals entsteht.
Die äussere und die mittlere Zellschicht der Retina werden durch die äussere plexiforme Schicht getrennt, dir dir Kontaktierungsstellen zwischen Photorezeptor-, Bipolar- und Horizontalzellen enthält.
Bipolarzellen sind direkt an eine oder mehrere Photorezeptorzellen gekoppelt, empfangen aber auf indlrektem Weg auch Signale von weiter entfernten Photorezeptorzellen. Der Bereich der Netzhaut, über den Lichtreize eine bestimmte Bipolarzelle beeinflussen, wird rezeptives Feld dieser Zell genannt. Es gibt zwei Typen dieser Zellart. Man nennt sie On-Zentrum- und Off-Zentrum-Zellen, je nachdem, ob sie depolarisiert (erregt) oder hyperpolarisiert (gehemmt) werden, wenn die direkt an sie gekoppelten Photorezeptorzellen beleuchtet werden. In beiden Fällen wirkt das indirekte Signal dem direkten entgegen, su dass eine On-Zentrum-Zelle erregt wird, wenn ein Lichtreiz auf die Mitte ihres rezeptiven Feldes projiziert wird, und gehemmt wird, wenn der Lichtreiz auf desson Peripherie fällt. Bei einer Off-Zentrum-Zelle ist das genau umgekehrt. Das rezeptive Feld einer Bipollarzelle besteht in der Fovea centralis aus einigen wenigen Photorezeptorzellen, wächst aber auf einen Durchmesser von bis zu zwei Millimetern in der Peripherie, wo sich die rezeptiven Felder benachbartrr Bipolarzellen also xu einem grossen Teil überlappen.
Horizontalzellen stellen über chjmische Synapsen laterale Verbindungen zwischen Photorezeptorzellen oder Bipolarzellen her, sind untereinander aber wie die Photorezeptoren über Gap junctions verbunden. Sie sind dünner gesät als Bipolarzellen und reagieren langsamer auf einen Reiz als Photorezeptorzellen. Wie die Photorezeptorzellen werden sie durch Stimulation hyperpolarisiert.
Da die rezeptiven Felder der Bipolar- und Horizontalzellen etwa gleich gross sind, wird angenommen, dass die Horizontalzellen mindestens teilweise für die Wechselwirkungen auf dem indirekten Weg verantwortlich sind. Photoreceptor-, Bipolar-und Horizontalzellen generieren keine Aktionspotentiale wie die meisten anderen Neuronen, sondern leiten ihre Signale durch analoge graduierte Potentialünderungen weiter. Die Signalstärke wird also in der Amplitude der Potentialänderung kodiert. Diese Methode funktieniert bei Nervenzellen nur über kleinere Strecken, da die Verluste und Zeitkconstanten sonst zu gross wurden. Die Länge der lateralen Verbindungen innerhalb der Netzhaut entspricht etwa der Grösse der rezeptiven Felder. Die Längsten Verbidungen sind also nur etwa einem Millimeter lang.
Zwishen der mittleren und der inneren Zellschicht der Retina liegt die innere plexiforme Schicht, in der Bipolar-, Amakrin und Ganglienzellen miteinander verbunden sind.
Die Ganglienzellen haben auch einen Zentrum-Umfeld-Aufbau und ähnliche rezeptive Felder wie die Bipolarzellen Ihre Polarität ist dabei die gleiche wie die der sic innervierenden Bipolarzellen. Neben On-Zentrum- und Off-Zentrum-Zellen existiert auch ein On-Off-Typus, der an On- und Off-Zeutrum-Bipolarzellen gekoppelt ist. Im Gegensatz zu Bipolarzellen erzeugen Ganglienzellen Aktionspotentiale die über den Sehnerv und das Chiasma opticum, wo Signale von beiden Augen zusammenkommen, zum nächsten Sehzentrum im Hirn weitergeleitet werden. Bei niedrigeren Wirbeltieren handelt es sich dabei nin das Tectum opticum, bei höheren um den Corpus geniculatum tief im Hirn, der seinerseits mit dem visuellen Cortex im Hinterkopf in Verbindung steht. Ganglienzellen zeigen imDunkeln eine spontane Aktivität von etwa fünf bis zehn Nervenimpulsen pro Sekunde.
Amakrinzellen stellen laterale Vebindungen zwischen Bipolar- und Ganglienzellen
her. Es gibt davon rnehr als zwandig Klassen, die verschiedene Wechselwirkungen
vermitteln. Eine Klasse stellt zum Beispiel erregende elektrische Kopplungen
zwischen Bipolarzellen her, eine andere bewirkt negative Rückkopplungen
auf Bipolarzellen, die dadurch kurz nach ihrer Aktivierung gehemmt werden,
also eine zeitliche Hochpasscharakteristik aufweisen. Je nach ihrem zeitlichen
Verhalten unterscheidet man zwei Typen von Ganglienzellen: Die einen zeigen
eine lang anhaldende Stimulationsantwort, die anderen eine transiente Antwort,
entweder wegen Kopplung
an Bipolarzellen mit Hochpasscharakteristîk oder wegen Hemmung
durch Amakrinzellen. Insgesamt gibt es etwa eine Million Ganglienzellen,
also etwa hundertmal weniger als Photorezeptorzellen. In der Fovea centralis
ist jeder Zapfen nur an eine Bipolarzelle direkt gekoppelt, die wiederum
auch nur eine Ganglienzellen innerviert, in der Peripherie kommen mehrere
Phetorezeptoren - hauptsächlich Stäbchen - auf eine Bipolarzelle
und mehrere Bipolarzellen auf eine Ganglienzelle. Dies erhöht die
Empfindlichkeit, reduziert aber das räumliche Auflösungsvermögen
der Ganglienzelle. Durch die antagonistische Wirkung ihres Zentrum-Umfeld-Aulbaus
reagieren Bipolar- und Ganglienzellen nur sehr schwach auf diffuses Licht,
das Zentrum und Umfeld gleichermassen stimuliert und somit bewirkt, dass
sich Erreguug und Hemmung die Waage halten.
Die Retina ist also trotz ihrer Lompaktheit und lokalen Struktur ein relativ komplexes System mit vielfältigen Verschaltungen, deren Zweck nicht unbedingt immer offensichtlich ist. Dennoch wurden verschiedene Funktionen durch intensive Forschung im Laufe der Zeit aufgeschlüsselt. Eine Auswahl davon wird im nächsten Abschnitt erläutert.
Funktion der Retina
Da wir in der Lage sind, unsere Augen in kurzer
Zeit fsst beliebig auszurichten, ermöglicht uns die gute Auflösung
und Farbdiskrimination in der Fovea centralis, die interessanten
Details in der Umgebung genau zu untersuchen, während die schlechter
auflösende und weitgehend monochromatische - dafür aber sehr
lichtempfindliche - Peripherie dazu dient, diese Details in einem grossen
Gesichtsfeld zu finden und unsere Augenbewegungen dorthin zu lenken.
Die logarithmische Charakteristik der Photorezeptorzellen bewirkt, dass die Potentialdifferenz, die ein lokaler Bildkontrast hervorruft, unabhängig von der Helligkeit ist. Diese Strategie trägt der Tatsache Rechnung, dass die meisten Objekte in unserer Umgebung Licht reflektieren oder streuen, das von einer global wirkenden Lichtquelle ausgesandt wird. Da das Streulicht im allgemeinen proportional zur Helligkeit der Lichtquelle ist, hängen die Verältnisse der Lichtstärken und somit die Kontraste nicht von der Beleuchtung ab. Gerade in natürlichen Umgebungen ist es wichtig, dass die Repräsentation der visuellen Signale weitgehend invariant gegenüber der Helligkeit der Lichtquelle ist, denn letzten Endes geht es darum, Objekte unabhängig von der Beleuchtung, die sich ja shon tagsüber um mehrere Grössnordnungen ändern kann, zu erkennen. Durch logaritmische Kompression wird ein riesiger Beleuchtungsstärkenbereich auf Potentialdifferenzen von weniger als hundert Millivolt übertragen. Damit eine Photorezeptorzelle aber trotzdem auf kleine Helligkeitsänderungen empfindlich ist, werden transiente Signale versäirkt und das Potential dann langsasm auf seinen Gleichgewichtswert zurückadaptiert. Auf dieser Adaptierungsfähigkeit basiert der enorm grosse Helligkeitsdynamikbereich der Photorezeptorzellen. Eine mesnchliche Photorezeptorzellen erreicht durch diese Strategie einen Helligkeitsdynamikbereich von fünf Grössenordnungen bei einer Kontrastauflösung von 2%, die weitgehend unabhängig von der Beleuchtung ist. Der Preis, den wir für diese Datenkodierung zahlen, ist unsere vergleichsweise schlecht ausgebildete Fähigkeit, absolute Helligkeiten abzuschA'tzen. So erscheint uns ein schwarzes Objekt im Sonnenlicht dunkler als ein weisses Objekt bei Zimmerbeleuchtung, auch wenn uns Lichtmessgeräte eines Besseren belehren (Bild 2).
![]()
Bild 2 Variables Helligkeitsempfinden
Wenn die Übergangslinie zwischen hellem und dunkIem Hintergrund
abgedeckt wird, scheint der einfarbige Ring Iinks heller zu sein als rechts.
Hinter der räumlichen Informationsverarbeitung stecken ähnliche Prinzipien wie hinter der zeitlichen. Der Bandpassfiltrer, der durch laterale Erregung und Hemmung implementiert wird, unterdrückt hohe Raumfrequenzen, um - auf Kosten der Auflösung - das Signal-Rausch-Verhältnis zu verbessern, und niedrige Raumfrequenzen, um das Signal zu dekorrelieren und langwellige Beleuchtungsvariationen herauszufiltern. So ergibt sich auch hier eine Kontrastkodierung, was im wesentlichen einer Kantenextraktion entspricht. Auch bei computergestützter Bildverarbeitung ist die Kantenextraktion normalerweise die erste und wichtigste Operation.
Die Auftrennung der Bipolar- und Ganglienzellen in On-Zentrum- und Off-Zentrum-Typen hat vermutlich damit zu tun, dass es für das Signal-Rausch-Verhältnis und den Energieverbrauch günstiger ist, wenn die Zellen im Grundtzustand weitgehend inaktiv sind, als wenn sie auf einem mittleren Aktivitatsniveau gehalten werden müssen, um positive und negative Signale kodieren zu können.
Die analogen gradruierten Potentiale, mit denen Potorezeptor- Bipolar- und Horizontalzellen Signale repräsentieren und kommunizieren, haben den Vorteil, dass sie kleine Werte besser kodieren als Aktionspotentiale dies tun und dass sie eine höhere Informationsübertragungsrate erlauben. Die wertkodierte Darstellung der visuellen Daten ermöglicht zudem - im Gegensatz zur zeitkodierten Repräsentation durch Nervenimpulse -die elegante Implementation einer Signalausbreitungsfunktion, die mit der Entfernung durch resistive Verbindungen graduell abnimmt, ohne dass die Zellen individuell weiträumig gekoppelt sein müssen.
Die Retina kann also in erster Näherung als nichlinearer zeitlicher und räumlicher Bandpassfilter für die Abbildung der Umwelt betrachtet werden. Ihre Funktion ist einerseits, im zentralen Bildbereich eine gute Auflösung und Farbisdiskrimination zu erreichen, und andererseits, das normalweise stark korrelierte Bild zu komprimieren, also die Bildinformation, die mehr in räumlichen und zeitlichen Veränderungen als in konstanten Graewerten liegt, effizienter zu kodieren. Dadurch kann die limitierte Datenhübertragungsbandbreite des Sehnervs besser genutzt und eine Überflutung der höheren Sehzentren im Gehirn durch redundante Eingangssignale verhindert werden
Implementationen künstlicher Retinae
Wie wir gesehen haben, benutzt die Netzhaut also eine Fülle von Strategien, um Bilddaten möglichst effizient aufzunehmen und zu kodieren. Die meisten davon können auch in künstlichen Systemen angewandt werden. Es sollte dabei aber nicht vergessen werden, dass solche System aus anderen Materialien aufgebaut sind und sich deswegen nicht alle Verarbeitungsstufen gleich gut emulieren lassen. Zudem sind auch nicht alle Elemente im Datenverarbeitungsprozess der Netzhaut hinreichend analysiert worden, abgesehen davron, dass gewisse Unterschiede zwischen den Retinae verschierdener Wirbeltierarten bestehen. So werden Netzhautemulationen immer einen gewissen Grad an Abstraktion und Vereienfachung beinhalten. Dabei ist es näturlich von Vorteil, Substrate zu wählen, bei denen physikalische Vorgänge ausgenutzt werden können, die ähnliche Charakteristika aufweisen wie die natürliche Retina.
Wir wollen im folgenden Möglichkeiten und Limitationen von Netzhautemulationen anhand einiger Systeme, die zur Entwicklung der künstlichen Retina wesentliche Beiträge geleistet haben, aufzeigen. Alle diese Systeme haben gemein, dass die Retina lediglich auf elektrischem Niveau emiliert wurde und kein Versuch unternommen wurde, biochemische Vorgänge nachzuvollziehen.
Ein frühes System [1] wurde aus diskreten elektrischen Standardkomponenten wie Photosensoren, Widerständen, Kondensatoren, Differentzverstärkern und Lampen zusammengebaut. Es enthielt 700 Photosensoren, die an zwei resistive Netzwerke angekoppelt waren, welche die erregende und hemmende laterale Wechselwirkung emilierten, die durch die Gap junctions der Photorezeptorzellen beziehungsweise der Horizontalzellen vermittelt wird (Bild 3).
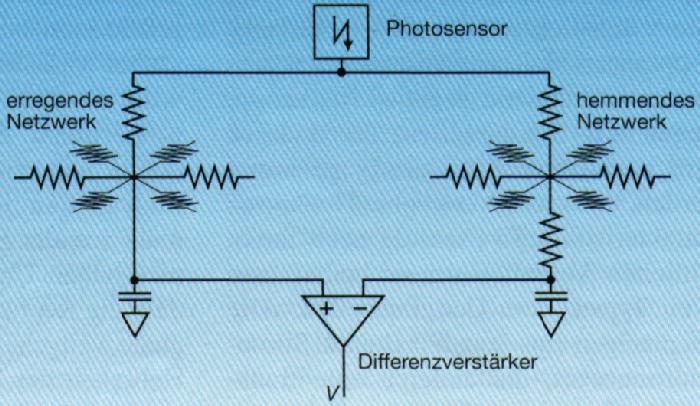
Bild 3 Schematischer Aufbau einer künstlichen Netzhaut [1]
Das Signal jedes Photosensors interagiert lateral über je ein
erregendes und ein hemmendes hexagonales Widerstandsnetzwerk mit denjenigen
(...) erregendes (...) Netzwerk
Die beiden Netzwerke waren als hexagonale Gitter (Bienenwabengeometrec) ausgelegt, wobei jeder Knoten nur mit seinen sechs nähsten Nachbarn direkt verbunden war. Der Einfluss des Potentials an einem Knoten auf anderen Knoten fällt bei einem solchen Netzwerk etwa exponentiell mit der Distanz ab. Die Widerstandsparameter waren dabei so gewählt, dass die Signale im erregenden Netzwerk stärker abfielen als im hemmenden, wie das beim biologischen Vorbild auch der Fall ist. Die Ansprechzeiten der beiden Netzwerke wurden durch Kondonsatoren an den entsprechenden Knoten bestimmt. Jedem Photorezeptor wurde je ein Knoten in den beiden Netzwerken zugeordnet. Die Signale dieser beiden Knoten wurden subtrahiert in einer Schaltung, die eine Bipolair- oder Ganglienzelle emuierte und deren Ausgangssignal eine Lampe ansteuerte. Auf der zweidimensionalen Lampenanzeige konnte dann eine Zentrum-Umfeld-Antwort beobachtet werden mit einer kürzeren Ansprechzeit für die Erregung als für die Hemmung, wie sie für solche Zellen typisch ist (Bild 4). Die räumliche Zentrim-Umfeld-Charakteristik wurde also durch die Differenz zweier radial etwa exponentiell abfallender Funktionen erzeugt.
![]()
Bild 4 Impulsantworten der kûnstîirhen Netzhaut von
Bild 2
a Interpolierte räumliche Verteilung des Ausgangssignals
entlang einer Achse bei Beleuchtung eines einzigen Photosensors
b Zeitlicher Verlauf des Ausgangssignals an einer Stelle
bei grossflächiger Stimulation mit einem kurzen Lichtblitz. Der Nulldurchgang
des Signals kommt durch die grössere Zeitkonstante des hemmenden Widerstandsnetzwerkes
zustande.
Durch die Verwendung von Widerstandsnetzwerken funktionierte das ganze System parallel und in Echtzeit ohne Iterarionen und Konvergenzprobleme, die mit Simulationen rückkoppelnder Schaltungen einhergehen würden, und vor allem - wie auch unser Hirn - ohne taktgebende Uhr, die ja das Herz eines heutigen Computers bildet.
Knapp zwanzig Jahre später wurde damit begonnen, Retina-Emulationen als integrierte Schaltungen herzustellen. Hierbei hat sich Silizium als sehr geeignetes Substrat erwiesen. Dies liegt zunächst einmal daran, dass die Siliziumtechnologie schon sehr ausgereift ist und man sich mittlerweile preisgünstigen Zugriff dazu verschaffen kann, ohne aufwendige Apparaturen kaufen zu müssen. Entscheidend ist aber auch, dass sich mit Standardsiliziumprozessen gute Photosensoren herstellen lassen, deren spektrale Empfindlichkeit derjenigen der Stäbchenzellen recht ähnlich ist. Diese Photosensoren lassen sich mit elektrischen Schaltungen auf eineim Siliziumchip integrieren. Integrierte schaltungen sind jedoch im wesentlichen zweidimensional, und so müssen die Schaltungselemente, die Zellen aus verschiedenen Schiechten der Retina entsprechen, in dieselbe Ebene geklappt werden (Bild 5).

Da die Nettzaut jedooh nur aus wenigen Zellschichten besteht und für die lateralen Wechselwirkungen nur nächste Nachbarn miteinander verdrahtet werden müssen, lässt sich dies - natürlich mit reduzierter Auflösung - durchaus bewerksrelligen. Neben ihren offensichtlichen Vorteilen wie ihrer mechanischen Robustheit, geringen Grösse und geringem Leistungsverbrauch haben integrierte Emulationen allerdings einen wesentlichen Nachteil.p.23Es handelt sich dabei um die Tatsache, dass sich bei kleinen Strukturen im allgemeinen und bei integrierten Analogschaltungen im besonderen die Inhomogenität des Fabrikationsprozesses bemerkbar macht und somit das Problem von systematischen räumlichen Unregelmässigkeiten und eventuell sogar Fehlstellen inhärent ist. Ein Problem, das auch von biologischen Verarbeitungsstrukturen gelöst werden muss. Es ist deshalb eine Herausforderung, beim Entwurf retinaler Schaltungen und Datenverarbeitungsalgorithmen Methoden zu finden, die gegenüber Variabilität der Schaltungselemente tolerant sind und die auch den Ausfall einzelner Elemecte verkraften können. Letzteres ist schon durch die lokale Kopplung und parallele Datenverarbetung einer künstlichen Netzhaut teilweise gewährleistet, decnn ein Schaltungsdefekt beeinstflusst dort nur eine Umgebung evon der Grösse des rezeptiven Feldes um den entsprechenden Bildpunkt. Bei rechnergestützter serieller Bildverarbeitung kann ein einzelner defekter Transistor unter Umständen den ganzen Verarbeitungsprozess lahmlegen.
Die erste integtierte Retina auf Siliziumbasis [2] besass Photorezeptorzellen mit logarithmischer Charakteristik über vier bis fünf Grössenordnungen in der Beleuchtungsstärke. Ein hexagonales Nettwerk von nichtlinearen regelbaren Widerständen mit relativ hohen Streukapazitäten übeitrug die hemmende Wirkung der Horizontazellen. In Bipolarzellen wurde dann das Potential eines Knotens von demjenigen des dazugehörigen Photorezeptors subtrahiert, so dass sich eine Zemtrum - Umfeld - Charakteristik ergab und zeitliches Hochpassverhalten für ausgedehnte Lichtreize. Durch das Fehlen eines erregeden Netzwerks war kein räumlicher Tiefpassfilter zur Rauschunterdrückung vorhanden und das Zentrum der räumlichen Charakteristik nur einen Bildpunkt gross. Infolge der logarithmischen Kompression des Helligkeitssignals wurde durch die Subtraktion in den Bipolarzellen der Logarithmus des Kontrastes kodiert, und das Ausgangssignal war somit in erster Näherung unabhängig von der Bildhelligkeit. Alle Schaltungselemente wurdce mit sehr kleinen Strömen betrieben, so dass die ganze Schaltung mit einer Auflösung von 48 x 48 Bildpunktce eine Leistung von nur einem Zehntel Milliwatt verbrauchte.
Dabei dieser Retina keine Signale im oben beschriebenen räumlichen Rauschen untergingen, benutzte eine spätere Fassung [3] einen Rückkopplungsmechanismus, durch den das System bei homogener Beleuchtung kalibriert werden konnte. Zur Speicherungg der Kalibration wurde eine Methode angewandt, die sonst für reprogrammierbare digitale Speicherchips benutzt wird, nämlich die Speicherung elektrischer Ladung auf einer unter normalen Bedingungen isolierten Leiterplatte, die kapazitiv das Verhalten einer Ausleseschaltung beeinflusst. Der Speicherinhalt kann durch gezieltes Anlegen von Spannungen oder durch Beleuchtung mit ultravioletter Strahlung verändert werden. In vorliegendem FaIl wurde die zweite Methode verwendet. Das räumnliche Rauschen konnte damit bei der kalibrierton Beleuchtungsstärke drastisch reduziert werden. Da aber auch der Verstarkungsfaktor der einzelnen Elemente statistischen Fluckuationen unterworfen war, musste die Kalibration bei wesentlichen Änderungen der Beleuchtungsstärke wiederholt werden.
In einem weiteren Iterationsschritt [4] wurde deshalb die Sîliziumretina mit einem adaptiven Photosensor ausgestattet, der eine sehr ähnliche Charakteristik aufwies wie biologische Photosensoren und durch einen gut kontrollierbaren transianten Verstärkungsmechanismus das Signal - Rausch - Verhältnis entschendend verbesserte. So konnte auch ohne Kalibration ein akzeptables Ausgangssignal generiert werden. Bild 6 zeigt das transiente Ausgangssignal eienr Weiterentwicklung dieser Retina.
![]()
Bild 6 Signalverteilungen am Ausgang einer integrierten künstlichen
Netzhaut mit 125 x 94 Bildpunkten in hexagonaler Anordnung
Durch die hemmende Wechselwirkung zwischen benachbarten Bildpunkten
und die Verstärkung zeitlicher Änderungen in den einzelnen Photosensoren
werden sich bewegende Konturen im Bild hervorgehoben.
Die oben beschriebene integrierte Emulation und ihre Modifikationen verwendeten Spannungen zur Repräsentation der Signale. Leider ist die Implementierung von einigermassen linearen Widerstandsnetzwerken für grössere Spannungssignale in Standardsiliziumprozessen recht aufwendig.
Das Problem liegt darin, dass in integrierten Schaltungen sehr kleine Ströme fliessen und deshalb hochohmige Widesrtände erforderlich sind, um vernünfttig Spannungsgradienten zu erzeugen. Ein Standardsiliziumprozess verfügt aber nur über niederohmige resistive Schichten, weshalb man sich zur Implementation von solchen Widerstandselementen mit relativ aufwendigen Transistorschaltungen behelfen mess. Ein System mit zwei hexagonalen Widerstandsnetzwerken erfordeit sechs Widerstandselementen pro Photosensor, wodurch die erforderliche Schaltung pro Bildpunkt natürlich relativ gröss wird. Wie sich aber herausstellte, kann ein regelbares, mehr oder weniger lineares resistives Netzwerk mit eicem einzigen Transistor pro Widerstandselement hergestellt werdenc, wenn Ströme anstelle von Spannungen zur Datenrepräsentation verwendet werden.
Basierend auf dieser Erkenntnis wurde in der Folge eine kompatie Emulation der Wechselwirkung von Photorezeptorund Bipolarzellen mit je einem erregendon und einem hemmenden hexagonalen Widerstandsnetzwerk implementiert [5], dessen Ausgangsströme eine Zentrum-Umgebung-Charakteristik aufwiesen, die den Kontrast relativ unabhängig von der Beleuchtunsstärke kodierten. Das zeitliche Verhalten der biologischen Rerina wurcle dabei jedoch nicht modelliert. Durch das Fehlen der transienten Verstärkung trat das räumliche Rauschen wieder vermehrt in Erscheinung. Es konnte jedoch durch Glättung des Signals über mehrere Photorezeptoren mittels errengender Intteraktion reduziert werden. Weil nur etwa zehn Transistoren pro Photorezeptor für die Bildverarbeitung benötigt wurden, konnten relativ grosse Netzwerke monolitisch integriert werden. Auf einem Chip mit einer Fläche von einem Quadratzentimeter wurde in einem konservativen Siliziumprozess eine Auflösung von bis zu 230x210 Bildpunkten erreicht. Mit einem modernen Prozess könnte die Auflösung um etwa einen Faktor drei pro Dimension gesteigert werden. Für Videoauflösung würde also weniger als oein Quadratzentimeter benötigt.
Wie bei der biologischen Netzhaut ist die verfügbare Datenübertragungsbandbreite bei integrierten Schaltungen beschränkt. Zwar sind die Schaltungen auf Siliziumchips etwa eine Million Mal schneller als die biologischen, dafür ist die Verdrahtung des Chips mit der Aussenwelt viel gröber als im Sehnerv, so dass nur einige wenige Kanäle zur Verfügung stehen. Die effiziente Bildkompression, die die Netzhaut durchführt, ist deswegen bei Emulationen mit Integrierten Schaltungen ebenso wichtig wie in der biologischen Netzhaut.
Spätere Versionen der obigen Schaltung [6] wurden deshalb mit einem zusätzlichen Elemente ausgestattet, das den Ausgangsstrom wie eine Gangliezelle in Aktionspotentiale mit Impulsfrequenzadaptierung umformte. Die Impulse aller Zellen wurden dann auf einen asynchronen digitalen Datenübertragungbus transferiert, wobei in jedem Impuls die Position der sendenden Zelle binär kodiert wurde, so dass ein Empfängersystem die Topographie der Zellstruktur rekonstruieren konnte. Andere Emulationen auf Siliziumbasis [7] konzentrierten sich auf die topographische Organisation der Netzhaut mit einer hochauflösenden Fovea centralis in der Mitte und radial nach aussen abnehmender Auflösung. Eine solche Topographie schränkt aberdlie Möglichkeiten zur Datenverarbeitung in einer zweidimensionalen Ebene stark ein, Weil in der Mitte zwischen den Photosensoren kaum für weitere Strukturen Platz ist. Eine solche Anordnung kann ausserdem ihr Potential nur in einem aktiven System voll entfalten, das durch Ausrichtung der optischen Achse des Abbildungsystems interessante Objekte auf die Fovea centralis projizieren kann. Wenn dies der Fall ist, können gewisse Bildverarbeitungsalgorithmen effizient implementiert werden wie zum Beispeil massstabsinvariante Objektrepräsentationen.
Die Aufnahme von Farbinformation durch drei verschiedene Typen von Zapfenzellen wird von herkömmlichen Videokameras, die mit einem Mosaik aus drei verschiedenen Farbfiltern versehen sind, bereits gut emuliert. Standardsiliziumprozesse können aufgrund der wellenlängenabhängigen Eindringtiefe der optischen Strahlung auch zur Aufnahme von Farbsignalen verwendet werden [8], allerdings mit relativ schlechter Diskriminationsfähigkeit.
Die in diesem Abschnitt beschriebenen Schaltungen bilden nur eine kleine Auswahl der künstlichen Systeme, die Aspekte der Bildverarbeitung durch die Netzhaut emulieren. Keines dieser Systeme implementiert alle bekannten wichtigen Funktionen der Rerina, gant zu schweigen von den Funktionen, die noch nicht ausreichend analysiert wurden. Welche Funktionen für eine Emulation wichtig sind, hängt von den anvisierten Anwendungen ab, die von der Modellierung biologiscger Prozesse bis zu industriellen Systemen reichen.
Anwendungen künstlicher Retinae
Durch die vielfähltigen Vorteile der Netzhaut gegenüber traditionellen
künstlichen Bildaufnahme- und - verarbeitungssystemen ergibt sich
eine grosse Palette von praktischen Anwendungen, bei denen es attraktiv
ist, solche System durch Netzhautemulation zu ersetzen. Dabei muss aber
den strukturellen und funktionalen Eigenheiten der künstlichen Retinae
auch bei der Konzeption des Gesamtsystems Rechnung getragen werden. So
eignen sie sich wegen ihrer limitierten Bildauftösung am besten zum
Einsatz in aktiven Systemen, von denen sic auf interessante Bildregionen
gerichtet werden können.
Ein interessantes Anwendungsgebiet wird einmal die Medizin werden, die versucht, degenerierte natürliche Retinae durch künstliche zu ersetzen. Diese Bestrebungen werden im vorliegenden Heft im Artikel von Alfred Stett und Hugo Hämmerle ab Seite 11 näher beleuchtet.
Viele Möglichkeiten eröffnen sich auch in der Robotik. Hier könnten künstliche Retinae in autonomen Systemen eingesetzt werden, die sich durch Lernprozesse eine Repräsentation ihrer Umwelt aufbauen.
Wie beim Menschen spielen dabei visuelle Eingangsdaten im allgemeinen eine zentrale Rolle. Künstliche Retinae sind für deren Erfassung geeignet, weil sie wichtige Anforderungen solcher Systeme erfüllen wie Echtzeitbildverarbeitung, Kompaktheit, Robustheit und niedriger Leistungsverbrauch. Ein illusttratives Beispeil für die Wichtigkeit dieser Anforderungen sind extraterrestriche Roboter. Man denke dabei an das Mars-Fahrzeug «Pathfinder», das aufgrund seiner ungenügenden Autonomie von der Erde und Mars verursachte lange Kommunicationszyklen und erlaubte daher nur langsame Bewegungen der Fahrzeuges. Hinzu kam ein grosser Energieverbrauch, der durch die relativ schwache Sonneneinstrahlung auf dem Mars nicht ausreichend durch Sonnenkollektoren abgedeckt werden konnte.
In der Verkehrstechnik interessiert sich vor allem die Automobilindustrie für die potentiellen Fähigkeiten der künstlichen Retinae. Hier sind unter anderem kostengünstige Produktionsmöglichkeiten gefragt. Da integrierte Retinachips im Gegensatz zu Videokamerachips mit billigen Standardsilizium prozessen hergestellt werden können, wäre diese Anforderung bei einer Massenproduktion zweifellos erfüllt. Der Hauptvorteil der Retienachips in einer solchen Anwendung ist aber die grosse Helligkeitsdynamik. Wenn eine Kamera zur Bildaufnahme für ein elektronisches System eingesetzt werden soll, das im Auto dem Fahrer unter anderem helfen soll, die Spur und den Abstand zum Vordermann zu halten und Verkehrszeichen zu erkennen, muss sie in der Lage sein, unter allen natürlichen Bedingungen gleichzeitig alle wichtigen Details zu erkennen. Unregelmässige Beleuchtungsverhältnisse, zum Beispiel bei Tunneldurchfahrten, Lichtreflexionen auf nasser Fahbahn oder bei Nachtfahrten dürfen die Funktion eines solchen Systems nicht Beeinträchtigen (Bild 7). Wie wir gesehen haben, könnte eine künstliche Retina mit solchen Problemen fertigwerden, während eine Videokamera allefalls durch Blendenregulierung mit globalen Helligkeitsvariationen zurechtkommt.

Bild 7 Signalverteilung arn Ausgang einer integrierten künstlichen
Netzhaut (vgl. Bild 5) bei qrosser
Helligkeitsdynamik in der optischen Abbildung
Ein grundlegendes Projekt unseres Institutes befasst sich mit der Emulatio verschiedener Teile des Cortex mit integrierten Schaltungen, welche übor einer asynchronen digitalen Datenbus durch Impulse kommunizieren [9] und sensorische Eingangssignale (z.B. von einer Retina) veraebeiten können. Im Rahmen eines solchen modularen Systems können biologische Datenverarbeitungs- und Lernprozesse studiert werden, die viellecht in nicht allzu ferner Zukunft auch ihren Weg in praktische Anwendungen finden werden. Wenn nur ein kleiner Teil der Leistungen des visuellen Cortex in einem solchen System repliziert werden könnte, wäre dies schon ein grosser Forschritt gegenüber existierenden künslichen Systemen. Allein schon ein Gesichtserkennungssystem, das ähnlich gut funktioniert wie unser biologisches. hätte ein riesiges Anwendungspotential.
Zusammenfassung
Wir haben gesehen, dass wir bei der Aujnahme und Verarbeitung vissueller
Information aus unserer Umgebung mit dem eigenen Sehsystem allen künstlichen
Systemen, die wir bisher gebaut hahen, in den meisten Hinsichten weit überlegen
sind. Bei der Konzeption künstlicher Systeme lohnt es sich deshalb,
biologische Lösungansätze in Betracht zu ziehen. Wie wir anhand
von künstlichen Retinae aufgezeigt haben, können biologisch inspirierte
Systeme effizient implementiert werden und sind in vielen Bereichen einsetzbar.
Adresse des Autors
Dr. Jörg Kramer, Institut für Neuroinformatik, Universität
un ETH
Zürich, Winterthurstrasse 190, 8057 Zürich
Literatur
[1] K. Fukushima, Y. Yamaguchi, M. Yasuda, S. Nagata: An Electronic
Model of the Retina. Proc. IEEE 58 (1970) 12, 1950-951.
[2] C.A. Mead, M-A. Mahowald: A Silicon Model of Early Visual
Processing. Neural Networks 1(1988), 91-97.
[3] CA. Mead: Adaptive Retina. In: Analog VLSI Implementation
of Neural Systems. Boston: Kluwer Academic 1989, pp. 239-246.
[4] M.A. Mahowald: Silicon Retina with Adaptive Photo receptors.
Proc. SPIE 1473 (1991), 52-58.
[5] K. A. Boahen, A. G. Andreou: A Contrast Sensitive Silicon
Retina with Reciprocal Synapses. In: Adv. Neural Information Processing
Systems 4. San Mateo, CA: Morgan Kaufmann 1992, pp. 764-772,
[6] K.A. Boahen: A Retinomorphic Vision System. IEEE Micro 16(19911)5,30-39.
(7] J. Van der Spieget G. Kreider, C. Claeys, I. Debusschere,
G. Sandint, P. Dario, F. Fantint, B. Belluti,
G. Soncini: A
Foveated Retina-like Sensor Using CCD Technology. Norwell, MA: Kluwer Academic
1989.
[8] P. Seitz; D. Leipold, J. Kramer; J.M. Raynor: Smart Optical
and Image Sensors Fabricated with Industrial CMOS/CCD Semiconductor Processes.
Proc. SPIE 1900(1993), 30-39.
[9] S. R. Deiss, R. J. Douglas, A. M. Whatley: A Pulse-coded
Communications infrastructure for Neuromorphic Systems. In: Pulsed Neural
Networks. Cambridge, MA: MIT 1999, pp. 157-178.